※独学の内容をまとめているので、間違っている部分があるかもしれません。詳しい方がおりましたら教えてください。m(_ _)m
- エマーソン効果とは
- 光合成のしくみについて【光化学系をまとめる前に】
- 光合成の明反応は光化学系(PhotoSystem : PS)Ⅰ、Ⅱ で行われる
- アンテナ蛋白質が光エネルギーを反応中心クロロフィルに渡す
- まとめ
- 参考
エマーソン効果とは
同じ量の 赤色光 と遠赤色光 を与えた時、遠赤色光 の光合成量は 赤色光 に比べてとても少なくなります。(700nm (遠赤色光の波長)を超えると極端に光合成量が低下することから、この現象は『レッドドロップ』と呼ばれています。)
しかし、 赤色光 + 遠赤色光 を与えると、各波長での光合成量を足した値よりも光合成量が増えます。

この現象は、ロバート・エマーソンが見つけたことからエマーソン(増強)効果と呼ばれています。
原著はこちら。
Red Drop and Role of Auxiliary Pigments in Photosynthesis(R. Emerson and E. Rabinowitch. 1960)
1960年代に発表してるようですが、当時はきっと設備も立派ではなかったでしょうから、凄いですねぇ。
でも、何故このような現象が起こるのでしょうか?
実は、二つの『光化学系』と呼ばれる色素 - 蛋白質複合体が存在するからなんです。
まずは光合成の大まかな仕組みからまとめていこうと思います。
光合成のしくみについて【光化学系をまとめる前に】
ものすごーーーく大雑把にいうと、光合成は
に分けられます。

チラコイドは拡大するとこのような構造になっています。

このチラコイドの膜に様々な蛋白質が存在していて、そこで明反応が行われています。


※各タンパク質はわざと大きく書いています。実際はもっと小さいのでご了承ください。また、実際にはもっとたくさんの種類の蛋白質が存在しています。
参考:L.テイツ / E.ザイガー テイツザイガー植物生理学(2017)培風館 p.173 - 201
チラコイド膜上に存在するタンパク質についてザッと載せましたが、明反応の主役こそが2つの色素 - 蛋白質複合体である、光化学系(PhotoSystem : PS)Ⅰ、Ⅱ です。
光合成の明反応は光化学系(PhotoSystem : PS)Ⅰ、Ⅱ で行われる
2つの光化学系はそれぞれ別の役割を持っています。
PSⅡ は
『光を受けて、水を酸素と水素イオンに分解し、電子をPSⅠ に渡す。』を行い、
PSⅠ は
『光を受けて、PSⅡ から送られてきた電子で NADP+ を還元』します。

最終的に作られた『NADPH』が暗反応のカルビン回路に使われます。
※ここでは省略していますが、チラコイド内膜に水素イオンが溜まるとストロマ側に出ようとします。その力を利用してATP合成酵素により、ATPが作られます。
それぞれの光化学系について、構造や組成をまとめてみます。
光化学系Ⅱ の構造と組成
PSⅡ の構造はこんな感じです。
光化学系Ⅱ を横から見たときの構造。画像上部はストロマ、下部はチラコイドの内側。
上画像は二量体の構造、下は単量体の構造
出典:光化学系Ⅱ - Wikipedia


光化学系Ⅱ を構成する色素や蛋白質はこんな感じで、反応に必要なタンパク質などで構成されています。

この中でも重要なのが P680 という特殊なクロロフィルです。P680 は他のクロロフィルと異なり、680 nm に光の吸収ピークがあり、光を受けると、光化学系Ⅰ へ電子を渡します。(不足した電子は水から受け取ります。)
ちなみに除草剤として使われるDCMU はプラストキノンに結合し、電子伝達を阻害するそうです。*1
光化学系Ⅰ の構造と組成
光化学系Ⅰ の構造はこんな感じ。
光化学系Ⅰ の構造。
参考:光化学系Ⅰ複合体の結晶構造解析

構成はこんな感じ。

参考:L.テイツ / E.ザイガー テイツザイガー植物生理学(2017)培風館 p.188、
光合成事典 - 光化学系Ⅰ複合体の結晶構造解析
こちらも重要なのは P700 という特殊なクロロフィル。700 nm に光の吸収ピークがあり、光を受けると、最終的にNADPへ電子を渡します。(不足した電子は光化学系Ⅱから受け取ります。)
ちなみに、除草剤として使われる「パラコート」は光化学系Ⅰの電子受容体から電子を受け取った後、酸素と直接反応して、葉緑体や脂質に有害なスーパーオキシドを作るそうです。*2
エマーソン効果はP680とP700の吸光ピークが異なるため起こる
ここまで書いてきたように、
となっております。
仮に680nm の光だけを与えた場合、光化学系Ⅱはとても活発に働こうとしますが、光化学系Ⅰ があまり働かないので、そこまで光合成が進みません。

光化学系Ⅱ は光化学系Ⅰ に電子を与えたい。でも光化学系Ⅰ は電子をフェレドキシンに電子を与えることができないので、光化学系Ⅱ から電子を受け取ることができません。
結果、明反応は進みません。
700nm の光を与えた時も同様、光化学系Ⅰ はとても活発ですが、光化学系Ⅱ が全く働かないので、光合成は進みません。

光化学系Ⅰ は電子を受け取りたいのに、光化学系Ⅱ から電子が送られてこない。。。
680, 700 nm の光を両方与えると、どちらの光化学系も活性化し、とても光合成の効率が上がります。

これがエマーソン効果のカラクリです。
光合成を効率よく行うには 680, 700 nm の両方の光が必要だということですね。
「じゃあ、他の光は使えないの?」
と思いますが、そんなことはないですよね。他波長の光でも光合成は進みます。最後に他波長の光をどのように光合成に利用しているのかについてまとめます。
アンテナ蛋白質が光エネルギーを反応中心クロロフィルに渡す
光化学系にはアンテナ蛋白質が結合している
チラコイド膜上には、アンテナ蛋白質と呼ばれる色素 - 蛋白質複合体が結合しており、光エネルギーを集める役割を持っています。代表的なアンテナ蛋白質はPSⅠ, Ⅱ に優先的に結合するLHCⅠ, Ⅱ (Light Harvesting Complex Ⅰ, Ⅱ)です。

LHCⅡ の構造はある程度明らかになっているようで、14 分子のクロロフィルa とb 、4 分子のカロテノイドを結合しています。ルテイン(カロテノイドの一種)、ネオザンチン、ビオラザンチンも結合しているようです。*3
LHCⅡ の構造
出典:Kiwako SAKABE , Noriyoshi SAKABE, 高等植物の集光性複合体 (LHC-II) の結晶構造解析とその周辺

アンテナ蛋白質に含まれる光合成色素が P680 や P700 に光エネルギーを渡しています。
光エネルギーは蛍光共鳴励起エネルギー移動により他の色素に移っていく
光合成色素はどのように光エネルギーを渡しているのでしょうか?
光エネルギーは、蛍光共鳴励起エネルギー移動(Fluorescence Resonance Energy Transfar : FRET) という機構により、P680 or P700 へ移動します。
FRET は音叉の共鳴に例えられます。
振動している音叉の近くにもう1つの音叉を置くと、その音叉も振動し始めるように、励起した光合成色素の近くに別の光合成色素があると、その光合成色素も励起します。


FRETについてもう少し掘り下げてみます。
たとえば、β - カロテンは 500 nm に吸光ピークを持ちます。500 nm の光(光子)は仮に『100』のエネルギーを持つとしましょう。β - カロテンが励起状態になるには『100』の光エネルギーが必要です。

β - カロテンに 500 nm の光子がぶつかると、励起状態(活性化状態)になります。

近くにあるクロロフィル b が FRET により、励起状態になります。ただ、クロロフィルb は『70』で励起状態になれるので、余分なエネルギーは熱として放出されます。

同じようにして、反応中心のP680 または P700 へと光エネルギーが渡されていきます。
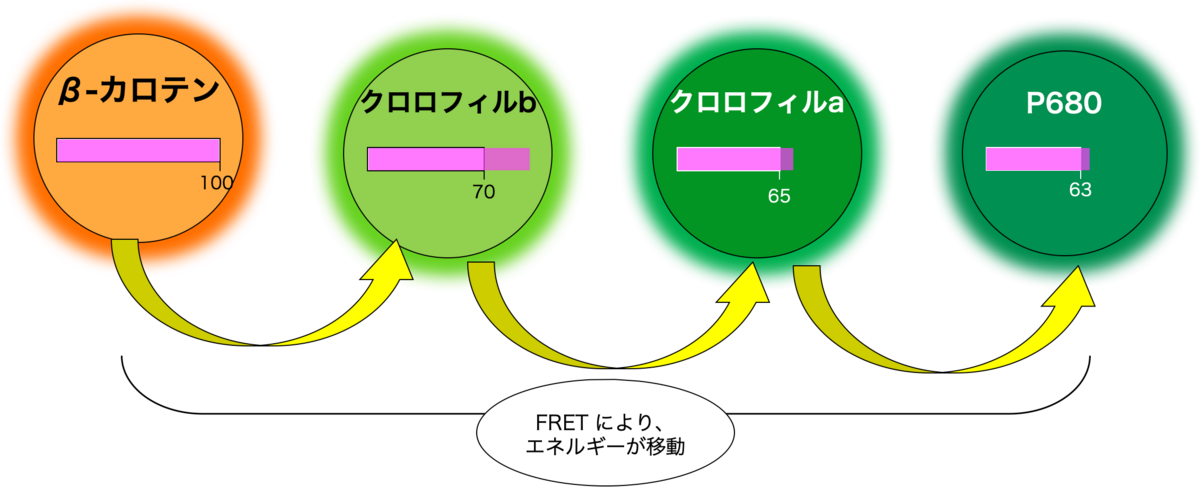
このように、どの波長の光が来ても、光エネルギーを反応中心に集められるような構造を取っています。
まとめ
以上、エマーソン効果と光合成の仕組みについてまとめてみました。これらを知ったからとて、栽培技術が劇的に向上するわけではないと思います。知識は場所をとりませんし、いつか役立つ時が来るかもしれません。(頑張ってまとめたので、役に立つ時が来るといいなぁ。)
参考
L.テイツ / E.ザイガー テイツザイガー植物生理学(2017)培風館
Red Drop and Role of Auxiliary Pigments in Photosynthesis(R. Emerson and E. Rabinowitch. 1960)